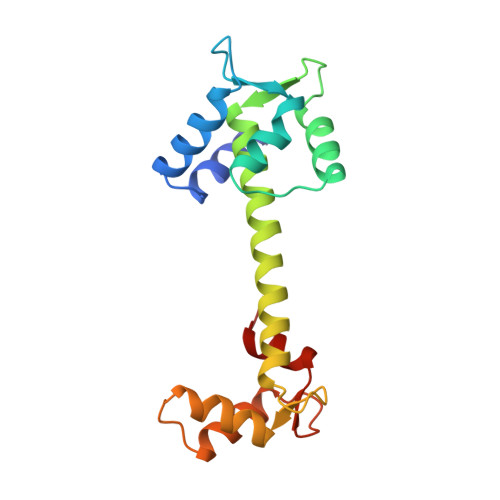
Refined crystal structure of troponin C from turkey skeletal muscle at 2.0 A resolution.
Herzberg, O., James, M.N.(1988) J Mol Biology 203: 761-779
- PubMed: 3210231 Search on PubMed
- DOI: https://doi.org/10.1016/0022-2836(88)90208-2
- Primary Citation Related Structures:
5TNC - PubMed Abstract:
The crystal structure of troponin C from turkey skeletal muscle has been refined at 2.0 A resolution (1 A = 0.1 nm). The resulting crystallographic R factor (R = sigma[[Fo[-[Fc[[/sigma[Fo[, where [Fo[ and [Fc[ are the observed and calculated structure factor amplitudes) is 0.155 for the 8054 reflections with intensities I greater than or equal to 2 sigma(I) within the 10 A to 2.0 A resolution range. With 66% of the residues in helical conformation, troponin C provides a good sample for helix analysis. The mean alpha-helix dihedral angles (phi, psi = -62 degrees, -42 degrees) agree with values observed for helical regions in other proteins. The helices are all curved and/or kinked. In particular, the 31 amino acid long inter-domain helix is smoothly curved, with a rather large radius of curvature of 137 A. Helix packing is different in the Ca2+-free domain (N-terminal) and the Ca2+-bound domain (C-terminal). The inter-helix angles for the two helix-loop-helix motifs in the regulatory domain are 133 degrees and 151 degrees, whereas the value for the two motifs in the C-terminal domain is 110 degrees, as observed in the EF-hands of parvalbumin. These differences affect the packing of the respective hydrophobic cores of each domain, in particular the disposition of aromatic rings. Pairwise arrangement of Ca2+-binding loops is common to both states, but the conformation is markedly different. Conversion of one to the other can be achieved by small cumulative changes of main-chain dihedral angles. The integrity of loop structure is maintained by numerous electrostatic interactions. Both salt bridges and carboxyl-carboxylate interactions are observed in TnC. There are more intramolecular (9) than intermolecular (1) salt bridges. Carboxyl-carboxylate interactions occur because the pH of the crystals is 5.0 and there is a multitude of aspartate and glutamate residues. One is intramolecular and four are intermolecular. Polar side-chain interactions occur more commonly with main-chain carbonyls and amides than with other polar side-chains. These interactions are mostly short range, and are similar to those observed in other proteins with one exception: negatively charged side-chains interact more frequently with main-chain carbonyl oxygen atoms. However, out of 19 such interactions, 10 involve oxygen atoms of the Ca2+ ligands. These unfavorable interactions are compensated by the favorable interactions with the Ca2+ ions and with main-chain amides. They are a trivial consequence of the tight fold of the Ca2+-binding loops.
- Medical Research Council of Canada Group, University of Alberta, Edmonton.
Organizational Affiliation: