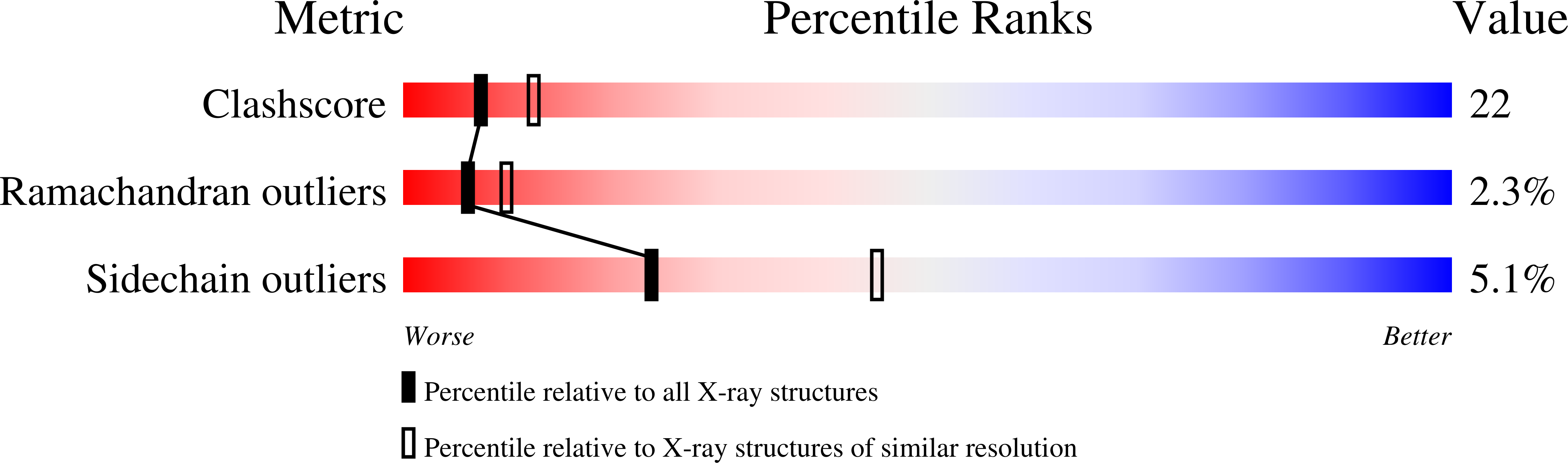
Complex of N-phosphonacetyl-L-aspartate with aspartate carbamoyltransferase. X-ray refinement, analysis of conformational changes and catalytic and allosteric mechanisms.
Ke, H.M., Lipscomb, W.N., Cho, Y.J., Honzatko, R.B.(1988) J Mol Biol 204: 725-747
- PubMed: 3066911
- DOI: https://doi.org/10.1016/0022-2836(88)90365-8
- Primary Citation of Related Structures:
8ATC - PubMed Abstract:
The allosteric enzyme aspartate carbamoyltransferase of Escherichia coli consists of six regulatory chains (R) and six catalytic chains (C) in D3 symmetry. The less active T conformation, complexed to the allosteric inhibitor CTP has been refined to 2.6 A (R-factor of 0.155). We now report refinement of the more active R conformation, complexed to the bisubstrate analog N-phosphonacetyl-L-aspartate (PALA) to 2.4 A (R-factor of 0.165, root-mean-square deviations from ideal bond distances and angles of 0.013 A and 2.2 degrees, respectively). The antiparallel beta-sheet in the revised segment 8-65 of the regulatory chain of the T conformation is confirmed in the R conformation, as is also the interchange of alanine 1 with the side-chain of asparagine 2 in the catalytic chain. The crystallographic asymmetric unit containing one-third of the molecule (C2R2) includes 925 sites for water molecules, and seven side-chains in alternative conformations. The gross conformational changes of the T to R transition are confirmed, including the elongation of the molecule along its threefold axis by 12 A, the relative reorientation of the catalytic trimers C3 by 10 degrees, and the rotation of the regulatory dimers R2 about the molecular twofold axis by 15 degrees. No changes occur in secondary structure. Essentially rigid-body transformations account for the movement of the four domains of each catalytic-regulatory unit; these include the allosteric effector domain, the equatorial (aspartate) domain, and the combination of the polar (carbamyl phosphate) and zinc domain, which moves as a rigid unit. However, interfaces change, for example the interface between the zinc domain of the R chain and the equatorial domain of the C chain, is nearly absent in the T state, but becomes extensive in the R state of the enzyme; also one catalytic-regulatory interface (C1-R4) of the T state disappears in the more active R state of the enzyme. Segments 50-55, 77-86 and 231-246 of the catalytic chain and segments 51-55, 67-72 and 150-153 of the regulatory chain show conformational changes that go beyond the rigid-body movement of their corresponding domains. The localized conformational changes in the catalytic chain all derive from the interactions of the enzyme with the inhibitor PALA; these changes may be important for the catalytic mechanism. The conformation changes in segments 67-72 and 150-153 of the regulatory chain may be important for the allosteric control of substrate binding. On the basis of the conformational differences of the T and R states of the enzyme, we present a plausible scheme for catalysis that assumes the ordered binding of substrates and the ordered release o
Organizational Affiliation:
Chemistry Department, Harvard University, Cambridge, MA 02138.